力学所在细胞粘附过程中受体-配体相互作用的研究中取得进展
细胞通过锚定在细胞膜上的受体蛋白与配体蛋白之间的相互作用与周围细胞发生粘附,进而调控诸如信号传导、免疫响应、组织形成以及癌症转移等关键细胞生命活动。因此,研究受体-配体相互作用是人们从分子尺度上认识细胞粘附及其调控机制最为关键的环节之一。
受体和配体之间的键合作用具有双向可逆性,其键合、解离反应的快慢与强弱,可分别由受体-配体反应动力学参数—正、逆反应速率以及亲和常数定量表征。与溶液中受体与配体之间的三维确定性反应不同,在细胞粘附过程中,受体-配体键合被限制在二维环境中,是一种受到外力调控的随机动力学过程。其键合的快慢、强弱以及寿命不仅与蛋白质固有性质有关,还会受到蛋白质-细胞膜相互作用的影响。
首先,细胞粘附过程中细胞膜自身存在热扰动。当膜间距在合适大小时,锚定在热扰动膜上的受体与配体才能发生键合。因此,受体-配体结合体将限制膜间距,并对细胞膜热扰动进行干扰。这会使得受体-配体结合体间产生相互吸引作用,并诱导粘附蛋白之间的协同键合。其物理图像如下:受体-配体结合体的形成会抑制细胞膜热扰动。受到抑制的细胞膜热扰动会反过来促进受体-配体键合以及额外结合体的形成。
其次,粘附蛋白通过与细胞膜相互作用可以诱导细胞膜曲率发生变化。诱导产生的膜曲率变化一方面会影响膜间距以及膜间相对粗糙度,另一方面会引起同层膜上蛋白质间顺式排斥作用。这两种因素会分别通过改变膜间距分布以及粘附蛋白质的有效平动面积影响受体-配体键合。
此外,细胞膜表面除了受体、配体蛋白外,还锚定着糖萼层。糖萼厚度一般大于受体-配体结合体的长度。这种尺寸差异会引起细胞膜的弹性变形并造成粘附蛋白的聚集,进而促进局部受体-配体结合体形成。同时,相比于粘附蛋白直径,糖萼聚合物横向尺寸较大,借助体积排空效应,同样会引起粘附蛋白的局部聚集,进而影响受体-配体键合。另外,糖萼层包含酸性粘多糖侧链,使其带有负电荷。该性质使得糖萼能够吸引并捕获周围液体内的阳离子,降低局部pH值,最终调控受体-配体键合。
最后,人们早期常采用由Singer与Nicholson 于1972 年提出的流动镶嵌模型来描述细胞膜结构。然而,累积证据表明,细胞膜并非均质体,而是含有许多富含鞘磷脂与胆固醇的微结构域。这些具有特定功能的微区常被称为“脂筏”。作为最重要的特性之一,脂筏通过对粘附蛋白的亲和力,导致受体、配体在脂筏内聚集。这种聚集不仅可以促进同层膜上蛋白质间的顺式相互作用,同时还会促进膜间受体-配体反式键合。其促进程度与脂筏面积分数、脂筏间吸引作用、脂筏弯曲刚度等脂筏特性密切相关。脂筏诱导的膜间受体-配体亲和常数的增加可归因于波动膜构象熵增加。
该工作从细胞膜热扰动、膜曲率、糖萼以及脂筏四个方面探究了蛋白质-细胞膜相互作用对膜间受体-配体键合的影响,深化了人们对蛋白质-细胞膜相互作用在细胞粘附过程中发挥作用的认识。
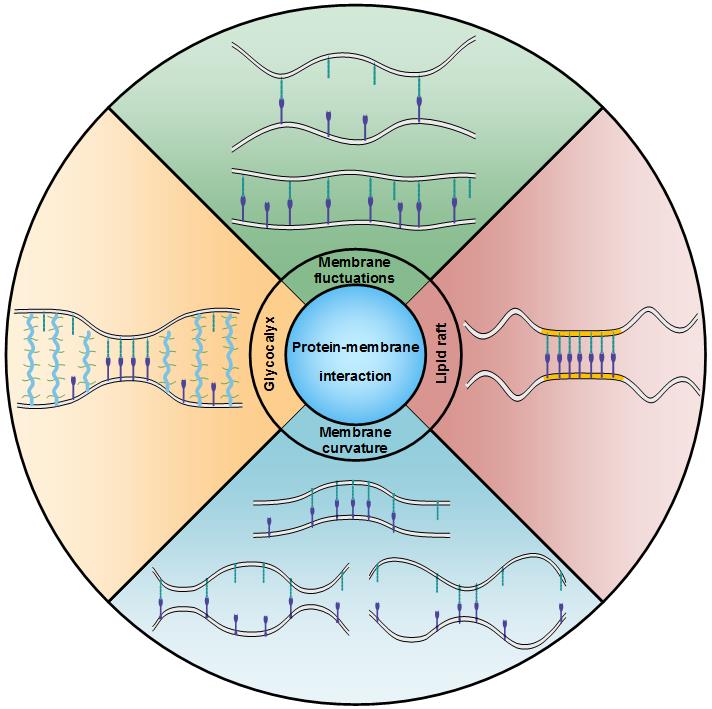
图1 细胞膜热扰动、膜曲率、糖萼以及脂筏共同调节膜间受体-配体相互作用
该工作以“Intercellular receptor-ligand binding: Effect of protein-membrane interaction”为题发表在分子生物学领域的TOP期刊Journal of Molecular Biology上。中国科学院力学研究所非线性力学国家重点实验室(LNM)李龙副研究员为文章第一作者。该研究工作得到国家自然科学基金与中国科学院青年创新促进会等项目资助。
附件下载:



